THE TENDON IS NOT A ROPE
Ask most people what a tendon is, and they’ll describe a rope. Something that connects muscle to bone, transmits force, and holds everything together. Strong, passive, unchanging. A bit of biological cabling.
It’s an understandable picture. Tendons are extraordinarily strong — the Achilles can withstand forces of up to 3,800 Newtons, several times your body weight with every running stride. They look like rope. They behave, mechanically, a little like rope. And for a long time, even in clinical thinking, they were treated like rope: something that either holds or snaps, with not much going on in between.
But that picture is wrong in the one way that matters most. A rope is inert. A tendon is alive.
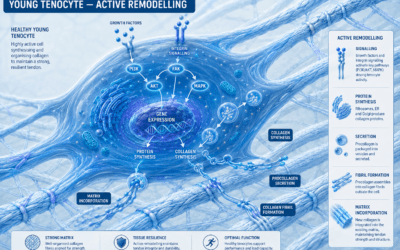
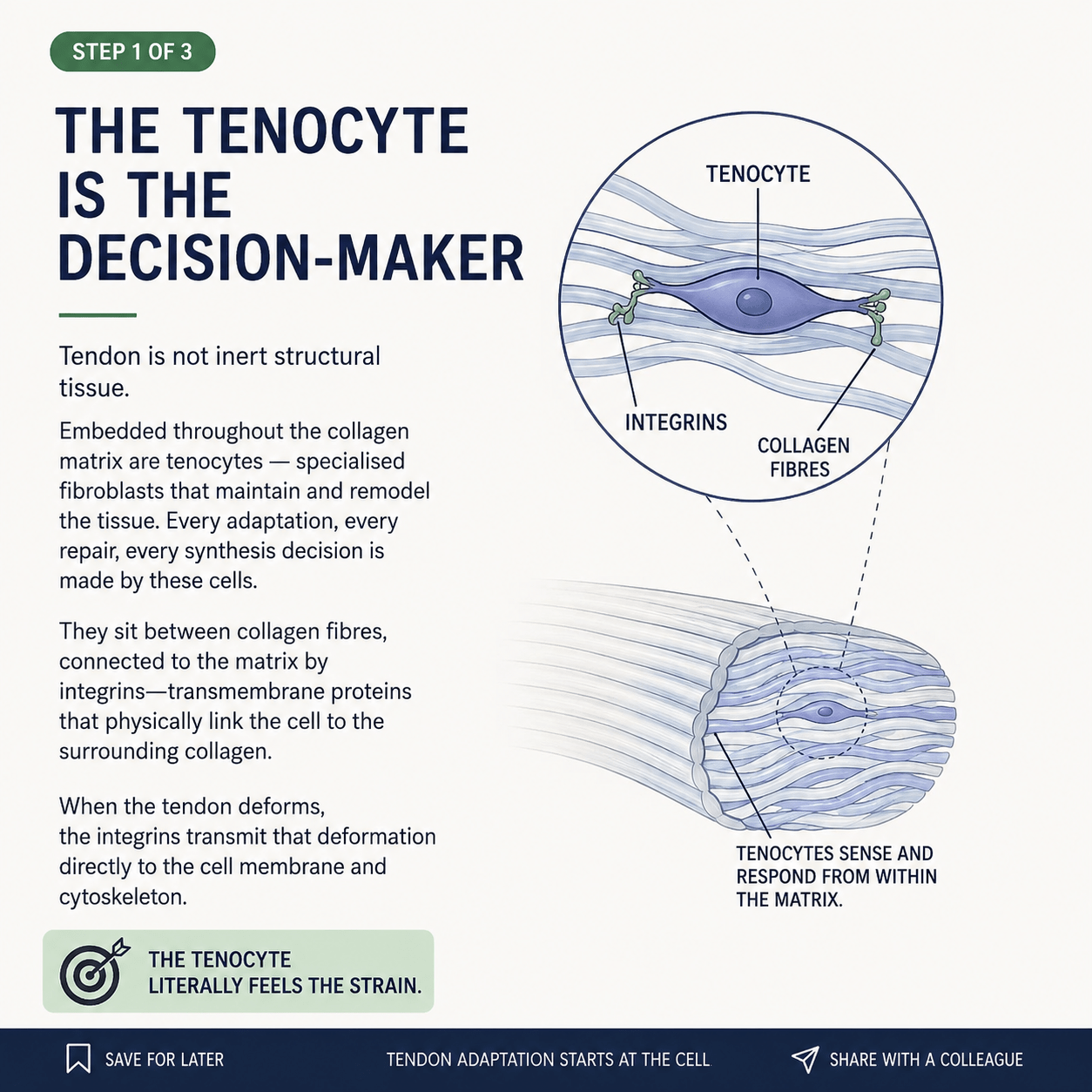
Embedded throughout the collagen matrix of every tendon are living cells called tenocytes — specialised fibroblasts that maintain, monitor, and remodel the tissue around them. They are not passengers. They are the decision-makers. Every repair, every adaptation, every decision to lay down new collagen is made by these cells. The strength of your tendon next year depends on the work these cells are doing right now.
And here is the elegant part. The tenocytes sit between the collagen fibres, physically tethered to the surrounding matrix by transmembrane proteins called integrins. Integrins are not just anchors — they are sensors. When the tendon deforms under load, the integrins transmit that deformation directly into the cell, into its membrane and its internal cytoskeleton.
The tenocyte literally feels the strain.
This changes how you have to think about everything. A rope doesn’t care whether you use it. A tendon does. When you load a tendon, you are not simply stressing a passive structure and hoping it holds — you are sending a signal to a living cell, which then decides whether to respond, how to respond, and how much new tissue to build. Loading is a conversation, not a stress test.
It also explains the things that puzzle people about tendons. Why they heal so slowly. Why rest alone never seems to fix them. Why some kinds of loading transform a stubborn tendon problem and others do nothing at all. Why a tendon can feel fine for months and then fail catastrophically in a single movement. None of that makes sense if a tendon is a rope. All of it makes sense once you understand it is a living, sensing, deciding tissue.
So if the tendon is not a rope — if it is a community of cells responding to the forces we put through them — then the obvious question becomes: what exactly are those cells responding to? What kind of loading do they listen to, and what kind do they ignore?
That is where we go next.
WHAT THE TENDON LISTENS TO
In the first post we established that a tendon is not a rope. It is a living tissue, full of cells called tenocytes that feel the strain we put through them and decide whether to respond. Loading is a conversation, not a stress test.
So the natural next question is: what makes the cell actually listen?
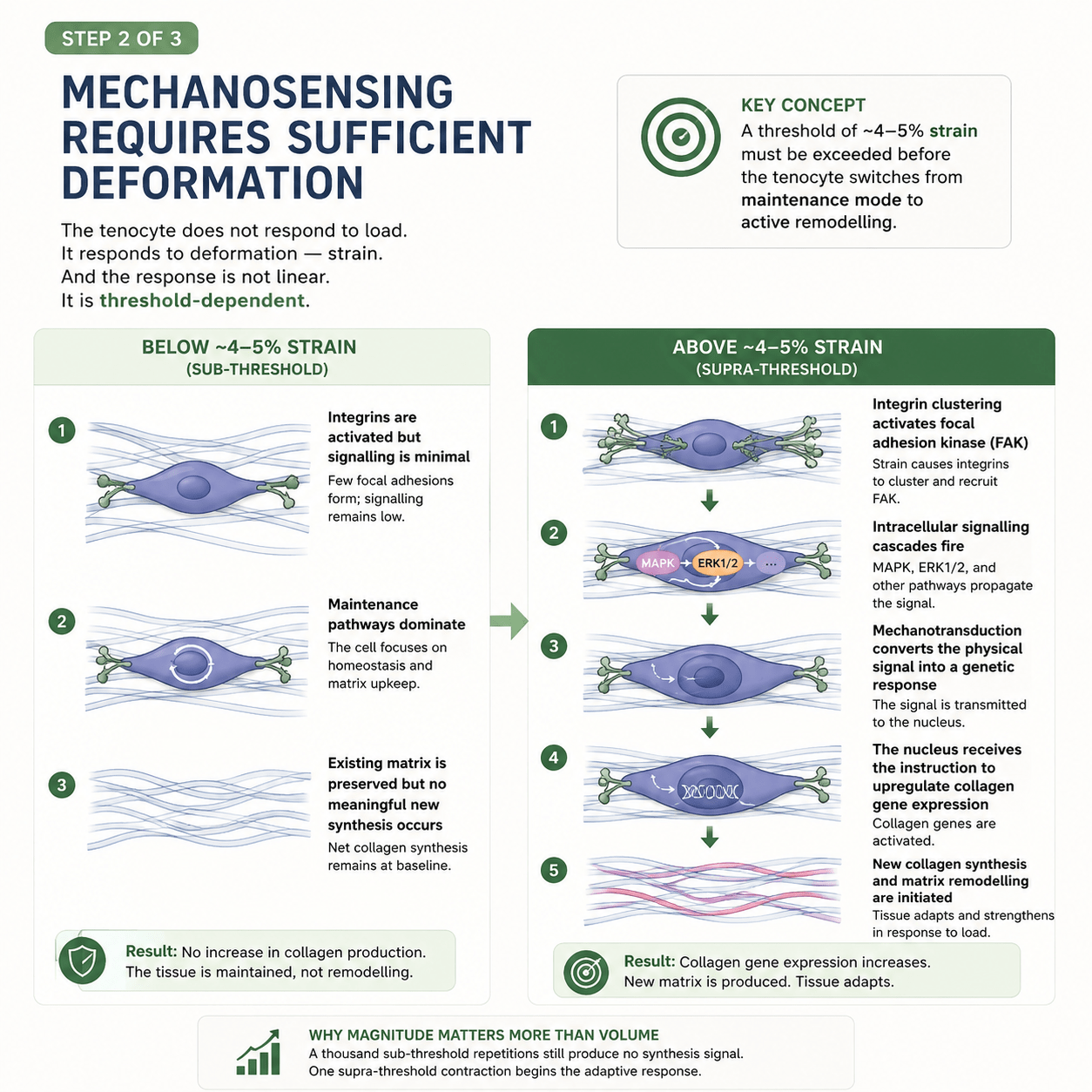
You might assume the answer is simply “load.” Use the tendon more, and it gets stronger. But that turns out to be wrong in an important way. The tenocyte does not respond to load in general. It responds to deformation — strain — and the response is not gradual. It is threshold-dependent.
Below a certain amount of strain, the cell essentially shrugs. The integrins that anchor it to the collagen are gently activated, but the signalling stays minimal. The cell stays in maintenance mode — keeping the existing tissue ticking over, repairing the ordinary wear of daily life, but building nothing new. You can load a tendon like this a thousand times and the net collagen synthesis barely moves. The tissue is maintained, not remodelled.
Then, at around 4 to 5% strain — which corresponds roughly to loads above 70% of the tendon’s maximum capacity — something changes. The threshold is crossed. The integrins cluster together and recruit an enzyme called focal adhesion kinase. Intracellular signalling cascades fire. The physical signal is converted into a genetic one and carried to the nucleus. The cell receives an instruction it did not receive a moment earlier: build new collagen. Synthesis begins. The matrix starts to remodel. The tissue adapts and strengthens.
This is the single most important idea in tendon rehabilitation, and it is the one most often missed.
It explains why magnitude matters more than volume. A thousand sub-threshold repetitions produce no synthesis signal at all. One contraction above the threshold begins the adaptive response. This is why walking, jogging, and ordinary daily movement — however much of it you do — will not heal a stubborn tendon. The load simply never gets high enough to switch the cell on.
It also explains why the advice to “just rest it” so often fails. Rest takes the load to zero. But zero is the deepest sub-threshold state of all. A rested tendon is not a remodelling tendon. It may calm down, the pain may settle, but the structural problem — the disorganised, under-built collagen — is not being addressed. The cell has not been given the one signal that would make it build.
And it explains why the right kind of loading can transform a tendon that years of rest, stretching, and anti-inflammatories never touched. Cross the threshold, repeatedly, with adequate recovery in between, and the cell does what it has been waiting to do all along.
So the threshold is the switch. Get above it, and the cell starts to build. Stay below it, and nothing changes no matter how hard you try.
But there is a second variable hiding in here — and it is the one almost nobody talks about. It is not enough to cross the threshold for an instant. How long you stay above it matters just as much as how hard you pull.
MAGNITUDE OPENS THE DOOR. TIME KEEPS IT OPEN.

we found the switch. At around 4 to 5% strain, the tenocyte crosses a threshold and begins to build new collagen. Below it, nothing meaningful happens. Above it, the adaptive response begins.
But crossing the threshold is only half the story. Because it turns out the cell does not just register that the threshold was crossed — it registers how long it stayed crossed. And that single fact changes everything about how a tendon should be loaded.
Think about two ways of loading a tendon to the same high force.
In the first, the movement is fast — one second up, one second down. The tendon shoots above the strain threshold, but only for a flicker. A few hundred milliseconds at the peak, and then it drops back down. The integrins cluster, signalling starts to fire — and then the stimulus is gone before the cellular machinery can fully engage. The signal is brief. The response is partial.
In the second, the movement is slow — three seconds up, three seconds down. The same tendon reaches the same force, but now it stays above the threshold for the full duration of the contraction. Six seconds, not a flicker. The integrin signalling sustains. Focal adhesion kinase keeps firing. The cascade has time to propagate all the way to the nucleus, and gene transcription is properly initiated. The signal is sustained. The response is complete.
Same load. Same threshold crossed. Completely different biological outcome.
This is why tempo matters as much as weight, and it is the part of tendon training that almost everyone gets wrong. The instinct, when we want a tendon to adapt, is to load it hard and fast — explosive, powerful, dynamic. But the tendon cell does not reward speed. It rewards time under tension. A fast, high-magnitude contraction opens the door for an instant. A slow contraction holds it open long enough for the cell to actually walk through.
It is worth being precise about what “slow” means here, because it is not vague. The protocol that the evidence supports is heavy slow resistance — loads above 70% of maximum, moved deliberately, typically three seconds in each direction, so that the tendon spends a meaningful stretch of every repetition above the strain threshold. Not ballistic. Not rushed. Heavy, and slow, and controlled.
It also explains a puzzle from elite sport. Plyometric training — jumping, bounding, explosive work — generates enormous tendon forces, often well above the threshold. So why does the research show it is comparatively poor at driving tendon adaptation? Because the loading duration per contraction is too brief. The forces are huge but the time above threshold is measured in milliseconds. The door is flung open and slammed shut before the cell can respond. The magnitude is there. The time is not.
And this is the heart of it. Magnitude opens the door. Time keeps it open.
If the strain threshold tells us how hard a tendon must be loaded to begin adapting, the time-under-tension principle tells us how that loading must be delivered. Heavy enough to cross the threshold. Slow enough to stay there. Repeated, with recovery, over weeks and months.
Which raises the last question in this sequence — the one about time on a much longer scale. We have talked about seconds within a single repetition. But tendon adaptation also unfolds over weeks and months, and there is a deep biological reason it cannot be rushed. The collagen a tendon is built from is, in a sense, older than most people realise.
Here’s the fourth blog post, picking up from the time-under-tension piece.
WHAT HAPPENS INSIDE THE CELL — AND WHY REST IS PART OF THE WORK
Gene expression and collagen synthesis
So far we have stayed at the level of forces and thresholds. The tendon is loaded, the cell feels the strain, and if the strain is high enough and held long enough, a switch is thrown. But what actually happens after that switch is thrown? What is the cell doing in the hours and days that follow?
This is where the timescale of tendon healing finally makes sense — and where one of the most counterintuitive principles in tendon training comes from: that rest is not the opposite of the work, it is part of the work.
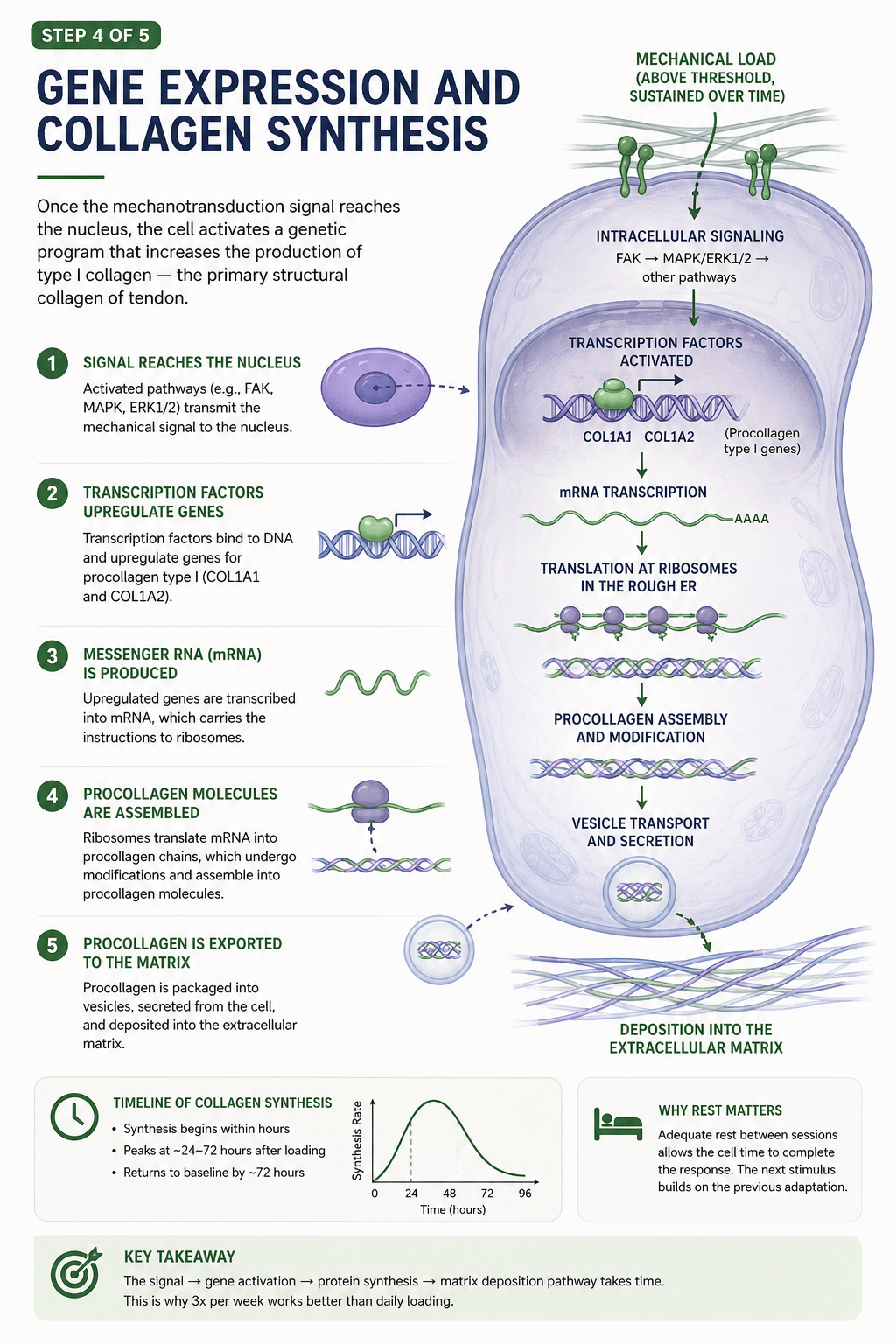
When mechanical load crosses the threshold and is sustained, the signal travels into the cell and reaches the nucleus. There, transcription factors switch on the genes for type I collagen — the primary structural collagen of tendon, encoded by genes called COL1A1 and COL1A2. Those genes are transcribed into messenger RNA. The mRNA carries the instructions out to the ribosomes, which translate them into procollagen chains. Those chains are assembled, modified, packaged into vesicles, and finally exported out of the cell into the surrounding matrix, where they will eventually become new tendon.
It is a beautiful, orderly production line. And crucially, it takes time.
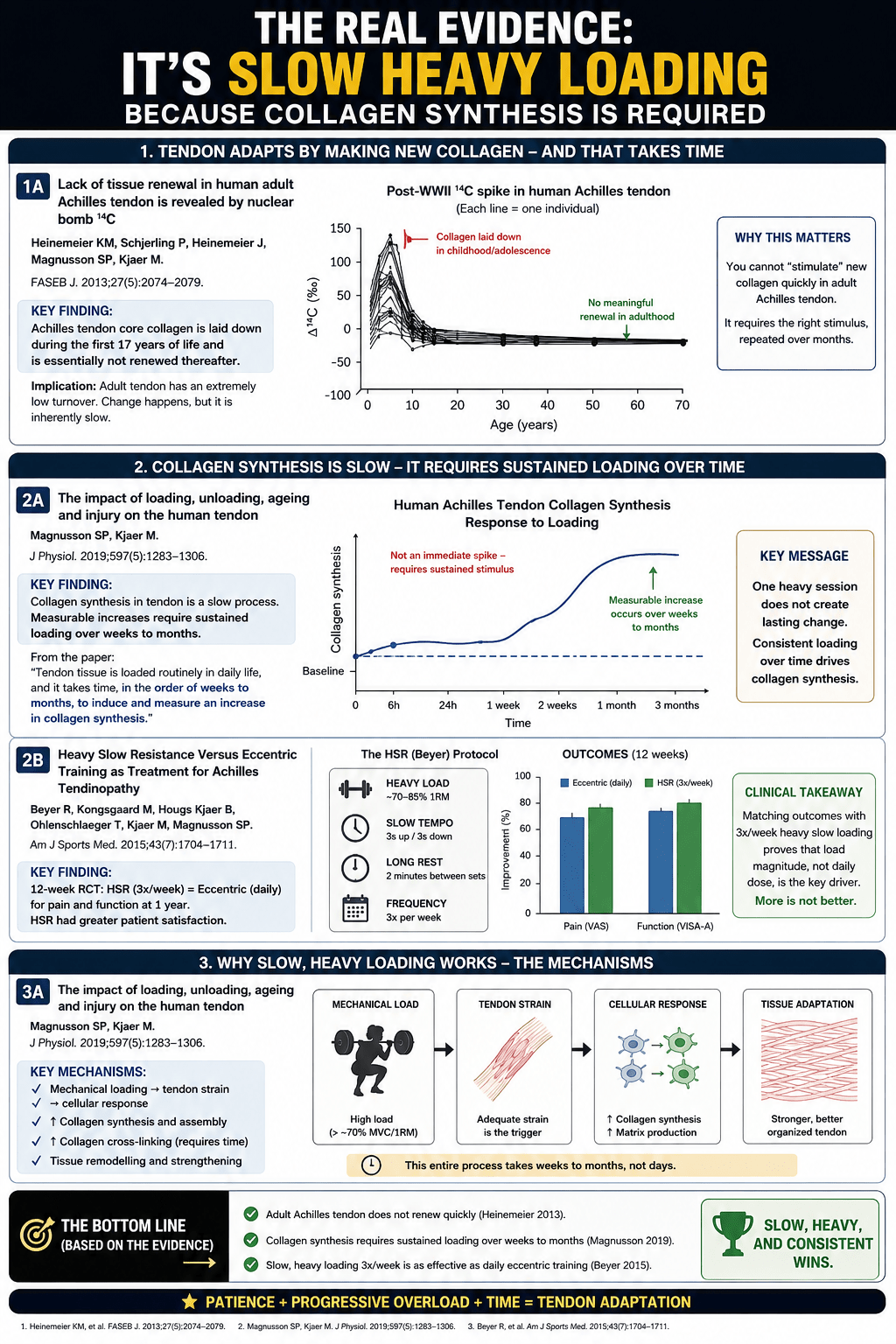
Collagen synthesis does not spike the instant you finish training. It begins within hours, rises steadily, and peaks somewhere around 24 to 72 hours after the loading session. Then it gradually returns to baseline by around the 72-hour mark. In other words, a single bout of heavy slow loading sets off a wave of new collagen production that unfolds over two to three days.
This one fact reorganises how you should think about training a tendon.
If the synthesis response to a single session lasts up to 72 hours, then loading the same tendon hard every single day does not give the cell time to finish what it started. You interrupt the production line before it has completed its run. You pile a new demand on a cell that is still working through the last one. More is not better — it is simply more interruption.
This is the biological reason that heavy slow resistance protocols are typically prescribed around three times per week, not daily. Three quality sessions, spaced to let each wave of synthesis run its course, drive more adaptation than seven rushed ones. The rest days are not time off from the work. They are when the work actually happens. The loading is the signal; the building takes place in the days that follow.
It also reframes what rest is for. When we tell someone to rest a tendon, we usually mean “stop loading it.” But the cell does not want zero load — we saw in the earlier posts that zero load is the deepest sub-threshold state of all, and produces no adaptation. What the cell wants is the right load, followed by enough recovery to complete the synthetic response, followed by the right load again. Rhythm, not absence. Stimulus, then space. Then stimulus again.
This is the quiet engine underneath every sensible tendon programme. Cross the threshold. Hold it long enough. Then step back and let the cell build. Repeat, patiently, with the days in between treated as part of the process rather than a pause in it.
And it points us to the final, deepest reason tendons cannot be rushed — one that goes beyond any single session or week. The collagen these cells are working with is not freshly made on demand. Most of the core collagen in your Achilles tendon has been there since before you were seventeen years old. What that means, and why it changes everything about how we treat tendons across a lifetime, is where we go next.
WHAT HAPPENS AFTER THE COLLAGEN IS MADE — AND WHY STRENGTH TAKES MONTHS, NOT DAYS
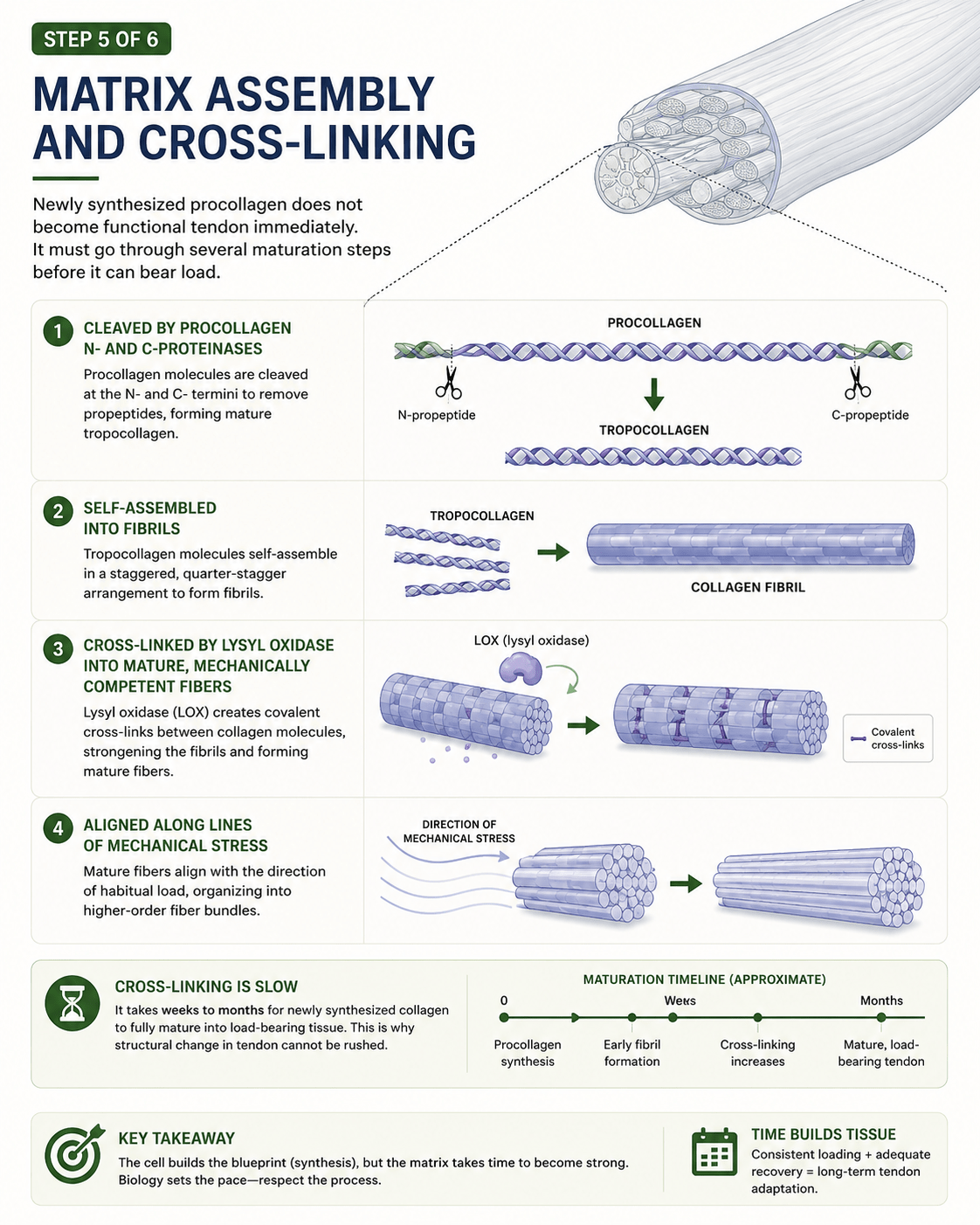
In the last image 4 we followed the production line all the way to its end: load crosses the threshold, the gene is switched on, the cell transcribes and translates and packages, and new procollagen is pushed out through the cell wall into the surrounding matrix. We left it there, with the synthesis wave rising over the two or three days that follow a session, because that is where most explanations stop. The collagen is made. Job done.
Except it isn’t. What leaves the cell is not tendon. Not yet.
This is the part of the story that almost never gets told, and it is the part that explains why tendons take so frustratingly long to get strong. Making the collagen is only the beginning. Turning that raw collagen into mature, load-bearing tissue is a separate process — slower, quieter, and largely invisible — and it unfolds not over hours but over weeks to months.
It happens in stages.
First, the new procollagen has to be trimmed. Inside the cell the molecule carries protective end-pieces, called propeptides, that keep it soluble and stop it assembling too soon. Once it is safely outside, specific enzymes — the procollagen N- and C-proteinases — snip those end-pieces off. What remains is tropocollagen: the true, finished building block of tendon.
Then those building blocks have to organise themselves. Tropocollagen molecules line up alongside one another in a precise, regularly staggered arrangement and pack together into fibrils. That regular offset is not cosmetic. It is the architecture that gives tendon its characteristic banded structure and its remarkable resistance to tensile load. The molecules are not thrown together; they are assembled.
But assembled is still not the same as strong. A freshly built fibril is organised yet mechanically weak — the molecules are sitting in the right places, but they are not yet bound to one another. The step that changes everything is cross-linking. An enzyme called lysyl oxidase forms covalent chemical bonds between adjacent collagen molecules, stitching the structure together from the inside. These cross-links are what convert neatly arranged collagen into mechanically competent fibre. Before they form, the tissue is fragile. After they form, it can bear load. And this is the slow step — the laying down and maturing of cross-links is measured in weeks and months, not days.
Finally, the maturing fibres align. They organise themselves along the lines of habitual load, so that the tendon is reinforced precisely in the direction it is being asked to work. The tissue builds itself in the shape of the demand placed on it.
Trim, assemble, cross-link, align. None of it is fast.
This is the missing timescale that makes sense of everything clinicians see. In the last post we learned that the synthesis response to a single session lasts up to seventy-two hours. That is the timescale of the raw material. But converting that raw material into strong tendon — cross-linked, aligned, mechanically competent — is a much longer job. It is why pain so often settles weeks before the tissue is actually ready, and why the most common mistake in tendon rehabilitation is stopping when it stops hurting. The symptoms can quieten in a few weeks. The building takes months.
It is also why the honest answer to “how long will this take?” is rarely the one people want to hear. We are not waiting for inflammation to calm down. We are waiting for biology to assemble, bond and align a new structure, one cross-link at a time, on a timescale it does not negotiate. Consistent loading, adequate recovery, and patience are not soft advice. They are the literal requirements of the process. The cell builds the blueprint, but the matrix takes time to become strong.
And there is one last layer beneath all of this. Every new fibre laid down here is being added to — and woven around — a core scaffold that is, for most of us, extraordinarily old. Most of the load-bearing collagen in your Achilles tendon has been there since before you were seventeen, and is barely renewed thereafter. What that means, and why it changes everything about how we treat tendons across a whole lifetime, is where we finish.
THE TENDON YOU BUILT BEFORE SEVENTEEN — AND WHY ALMOST NOTHING BUT SLOW, HEAVY LOADING WILL CHANGE IT
We have come, at last, to the fact that reframes all the others.
Across these posts we have watched the tendon behave like living tissue: sensing load, switching on genes, synthesising collagen over days, assembling and cross-linking it into strength over months. Everything so far has described building. But there is a hard limit on what can be built, and it sits underneath the whole story.
In 2013, Heinemeier and colleagues did something ingenious. They used the carbon-14 released into the atmosphere by post-war nuclear bomb testing as a biological timestamp, and traced it through human tendon tissue. What they found was sobering. The core collagen of the adult Achilles tendon is laid down before roughly the age of seventeen, and is essentially not renewed thereafter. The scaffold you finished building as a teenager is, in large part, the scaffold you carry for the rest of your life.
This changes the meaning of everything we have discussed. The synthesis we can drive with loading, and the cross-linking that matures over months, act mostly at the surface and the margins of that core. We are not constantly rebuilding the tendon from scratch. We are maintaining, reinforcing and slowly remodelling a structure that is, at its heart, decades old.
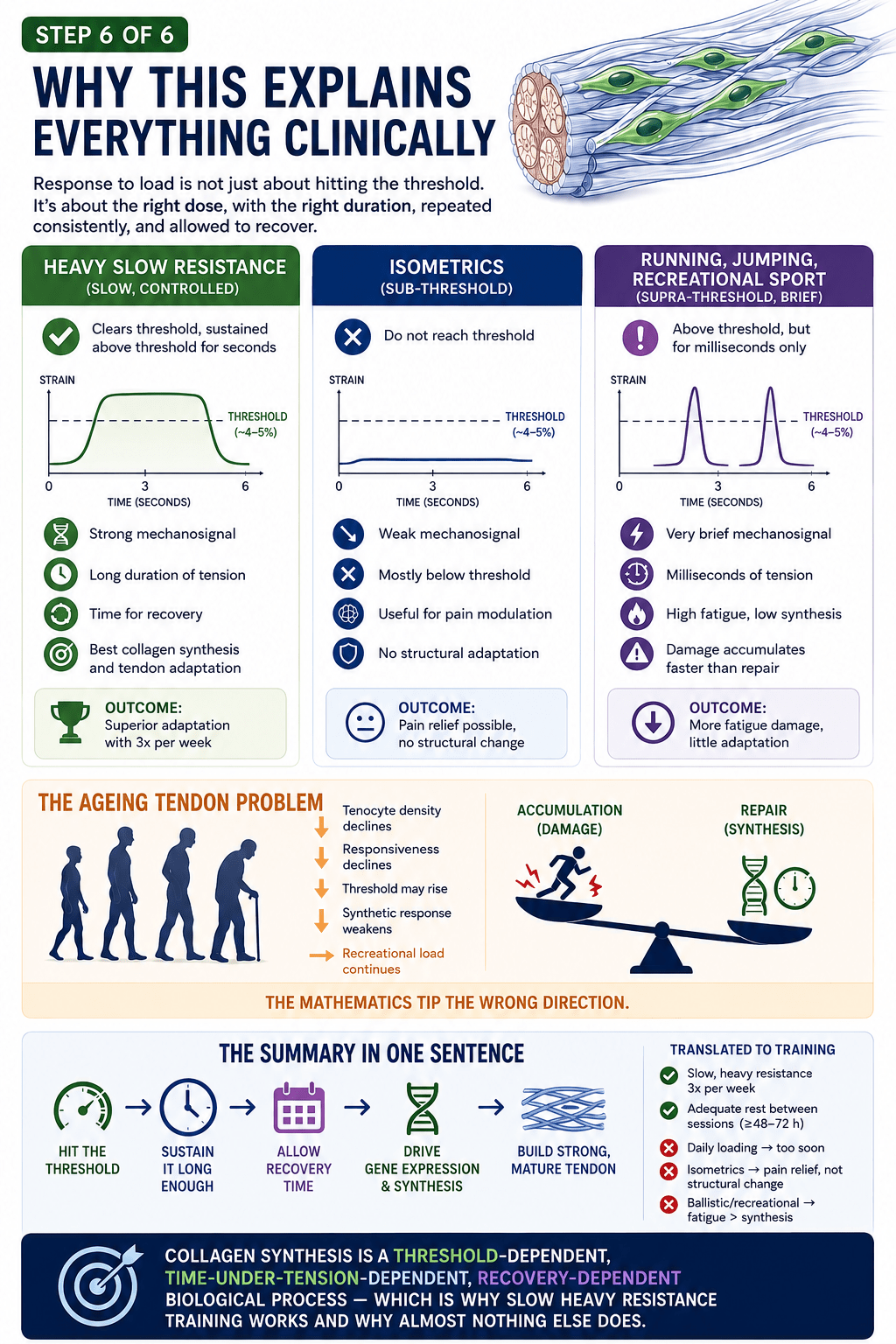
And here is where age turns the screw. As tendons age, the cells that do the maintenance work — the tenocytes — decline in number. Those that remain become less responsive to mechanical signals. The strain threshold needed to switch them on may rise, so the same load produces a smaller adaptive response. The synthetic machinery slows. Yet the loads of daily life, sport and recreation continue much as before. Damage accumulates on one side of the scales; repair struggles to keep up on the other. The mathematics tip, quietly, in the wrong direction.
Put together, these two facts — an ancient core that cannot be quickly replaced, and an ageing cell population that responds more weakly each decade — explain why tendons are so unforgiving, and why getting the loading right matters more, not less, as the years go on.
So what actually works?
This is where everything we have learned becomes a single, practical filter. Collagen synthesis is threshold-dependent: the strain has to be high enough, somewhere around four to five per cent, which in practice means loads above roughly seventy per cent of maximum (Magnusson and Kjaer, 2019). It is time-under-tension-dependent: the strain has to be held long enough for the cell to register it. And it is recovery-dependent: the synthetic response must be allowed to run its course over the days that follow. Hold those three requirements up against the things people actually do with their tendons, and the picture becomes strikingly clear.
Heavy slow resistance loading clears the threshold and holds it there for seconds at a time. Strong signal, long duration, and time to recover between sessions. This is the one common form of loading that consistently drives meaningful structural adaptation — and Bohm and colleagues’ meta-analysis confirms it does so regardless of whether the contraction is concentric, eccentric or isometric, provided the intensity is high enough and the work is sustained over twelve weeks or more.
Isometrics, held lightly, often sit below that structural threshold. They can be genuinely useful for calming pain, and they have a real place as an entry point for an irritable tendon. But pain relief is not the same thing as structural change, and we should be careful not to confuse the two.
And running, jumping and recreational sport? They send the tendon briefly above threshold — but only for milliseconds at a time. The signal is too short to drive much synthesis, while the cumulative fatigue and microdamage are very real. For a healthy, well-prepared tendon, this is simply sport. For a tendon already losing the race between damage and repair, it is load without the building stimulus to match it.
Which brings the whole series to its single, unsentimental conclusion. Collagen synthesis is a threshold-dependent, time-under-tension-dependent, recovery-dependent biological process. That is precisely why slow, heavy resistance training works — and why almost nothing else does. Not because it is fashionable, and not because eccentrics or any one protocol hold special magic, but because it is the only common form of loading that satisfies all three of biology’s conditions at once.
The tendon is not a rope. It is living tissue, built on an ancient scaffold, maintained by ageing cells, and changed only by the right load, held long enough, repeated patiently, with time to recover in between. Respect that, and the tissue will adapt. Ignore it, and no amount of effort will substitute for the one thing the tendon actually asks of us: the right work, done consistently, over time.
REFERENCES — TENDON BIOLOGY & ACHILLES RUPTURE SERIES
“Free full text” means the complete paper is openly available; “abstract” means the journal is subscription-only and the link opens the abstract and citation.
–
CORE PAPERS
1. Heinemeier KM, Schjerling P, Heinemeier J, Magnusson SP, Kjaer M. Lack of tissue renewal in human adult Achilles tendon is revealed by nuclear bomb ¹⁴C. FASEB J. 2013;27(5):2074–2079.
The carbon-14 study showing the core collagen is laid down before about age 17 and essentially not renewed thereafter.
Free full text: <https://pmc.ncbi.nlm.nih.gov/articles/PMC3633810/>
DOI: <https://doi.org/10.1096/fj.12-225599> · PMID: 23401563
1. Magnusson SP, Kjaer M. The impact of loading, unloading, ageing and injury on the human tendon. J Physiol. 2019;597(5):1283–1298.
The review tying together the strain threshold, ageing effects, and adaptation mechanisms.
Free full text: <https://pmc.ncbi.nlm.nih.gov/articles/PMC6395417/>
DOI: <https://doi.org/10.1113/JP275450> · PMID: 29920664
1. Bohm S, Mersmann F, Arampatzis A. Human tendon adaptation in response to mechanical loading: a systematic review and meta-analysis of exercise intervention studies on healthy adults. Sports Med Open. 2015;1:7.
The meta-analysis showing loading above ~70% of maximum drives stiffness gains, contraction type doesn’t matter, and 12+ weeks works best.
Free full text (open access): <https://doi.org/10.1186/s40798-015-0009-9>
Also at: <https://pmc.ncbi.nlm.nih.gov/articles/PMC5005577/>
1. Hoenig T, Gronwald T, Hollander K, Klein C, Frosch K-H, Ueblacker P, Rolvien T. Video analysis of Achilles tendon ruptures in professional male football (soccer) reveals underlying injury patterns and provides strategies for injury prevention. Knee Surg Sports Traumatol Arthrosc. 2023;31(6):2236–2245.
The video analysis of 80 ruptures: 94% indirect or non-contact, the five movement patterns, mean age 28.6, median return 202 days.
Free full text: <https://pmc.ncbi.nlm.nih.gov/articles/PMC10183418/>
DOI: <https://doi.org/10.1007/s00167-023-07384-1> · PMID: 36977780
1. Beyer R, Kongsgaard M, Hougs Kjær B, Øhlenschlæger T, Kjær M, Magnusson SP. Heavy slow resistance versus eccentric training as treatment for Achilles tendinopathy: a randomized controlled trial. Am J Sports Med. 2015;43(7):1704–1711.
The RCT showing heavy slow resistance and eccentric training give equally good results — supporting the point that intensity matters more than contraction type.
Abstract: <https://pubmed.ncbi.nlm.nih.gov/26018970/>
DOI: <https://doi.org/10.1177/0363546515584760>
1. Briggs-Price S, O’Neill S, Houchen-Wolloff L, et al. Incidence, demographics, characteristics and management of acute Achilles tendon rupture: an epidemiological study. PLoS One. 2024;19(6):e0304197.
The UK study reporting that 46.2% of rupture patients had prior Achilles pain or injury (of those, 61.1% on the ruptured side or bilaterally).
Free full text (open access): <https://pmc.ncbi.nlm.nih.gov/articles/PMC11192343/>
DOI: <https://doi.org/10.1371/journal.pone.0304197>
—
SUPPORTING EVIDENCE
1. Rio E, Kidgell D, Purdam C, Gaida J, Moseley GL, Pearce AJ, Cook J. Isometric exercise induces analgesia and reduces inhibition in patellar tendinopathy. Br J Sports Med. 2015;49(19):1277–1283.
The study behind the “isometrics for pain relief, not structural change” point (patellar tendon, but the analgesia principle is the relevant one).
Abstract: <https://pubmed.ncbi.nlm.nih.gov/25979840/>
DOI: <https://doi.org/10.1136/bjsports-2014-094386>